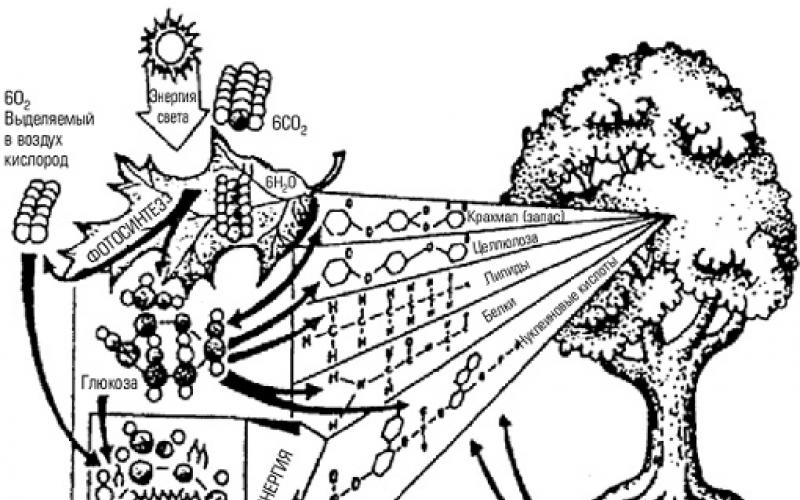
«... два американских учёных, Роберт МакАртур и Эдвард Уилсон , создали теорию R-К отбора. Теорию двух разных стратегий размножения живых существ.
Теория двух стратегий оказалась настолько удачной, что используется в ряде наук, признана практически всеми, вошла в учебники и в учебные пособия.
R-стратегия - это рождение за единицу времени как можно большего числа детёнышей.
О каждом из них можно практически не заботиться, и каждый детёныш имеет не очень много шансов выжить. Муха откладывает 5 млн. яичек - и что, она очень волнуется по поводу судьбы этих 5 млн. будущих маленьких мушек? Сотнями тысяч и миллионами откладывают яички насекомые, ракообразные, моллюски. Рыбы, которые вымётывают «всего» десятки тысяч икринок, тем более лягушки, вымётывающие тысячи икринок, - это просто идеальные родители в сравнении с более простыми существами. Конечно, и они никак не заботятся о своих потомках, но эти более сложные животные вынуждены выметывать более сложные, более крупные икринки - а тем самым выметывать меньшее число этих икринок. Некоторые виды рыб уже стараются охранять своих вылупившихся рыбят: строят им гнезда, нападают на появившихся хищников. Некоторые виды даже держат мальков в собственном рту, и там мальки спасаются в случае опасности.
Это уже элементы К-стратегии: рождения небольшого числа детёнышей, каждый из которых важен и ценен. Чем сложнее вид - тем ценнее для него каждая отдельная жизнь, тем меньше детёнышей погибает между рождением и смертью. Чем проще устроено живое существо, тем меньше его надо учить и готовить к жизни, тем быстрее оно становится взрослым.
Мышь может родить три раза в год по десять мышат. Роды у мыши проходят очень легко, а малыши через три недели становятся взрослыми. Они уже могут о себе сами позаботиться, мать выгоняет их и готова родить новых. Если мышата не будут погибать - мир скоро заполнится полчищами взрослых мышей. У более сложных животных - слонов, шимпанзе, лосей, зубров - детёнышей рождается меньше, и они реже погибают.
Но и у крупных сложных животных физиологическая норма - смертность 60-70% новорожденных. Самка шимпанзе и слона рожает за свою жизнь 10-15 раз. 7, 10 или даже 12 из этих детёнышей умрут до того, как станут взрослыми. Вырастут и сами дадут племя те самые 2 или 3 детёныша, которые необходимы для воспроизводства вида.
После катастроф при взрывах вулканов, после цунами новые острова и побережья «захватывают» живые существа с R-стратегией. Но вскоре более крупные, сложно устроенные животные с К-стратегией начинают господствовать. Эволюция - во многом борьба не за выживание, а за господство».
Буровский А.М., Феномен мозга. Тайны 100 миллиардов нейронов, М., «Яуза»; «Эксмо», 2010 г., с. 77-79.
В 1967 г. Р.Макартур и Е.Уилсон, анализируя динамику численности популяций, предложили r- и К-коэффициенты [ MacArtur R.H., Wilson E.O., 1967 ]. Мы не будем рассматривать их математический смысл, а используем эти коэффициенты для обозначения двух стратегий эволюционного развития живых существ.
r-Стратегия предполагает бурное размножение и короткую продолжительность жизни особей, а k-стратегия - низкий темп размножения и долгую жизнь. В соответствии с r-стратегией популяция развивается на переломных этапах своей истории, при изменении внешней среды, что способствует появлению новых признаков и захвату новых ареалов. К-стратегия характерна для благоденствия популяции в уже захваченном ареале и при сравнительно стабильных условиях. Очевидно, что у популяции вероятность новаций будет тем выше, чем она быстрее размножается и чем чаще происходит смена поколений, т.е. короче продолжительность жизни особей. Чтобы решить проблему переходных форм, r-стратегии недостаточно, желательно дополнить ее еще одним свойством, а именно повышенной жизнеспособностью, или лучшими качествами в борьбе за существование, на коротком (в сравнении с К-стратегией) временном промежутке, отведенном природой для жизни особи. Это в общем-то логично: за повышение жизнеспособности, как и за плодовитость, приходится платить, а плата эта - сокращение продолжительности жизни. Если жизнеспособность особей при r-стратегии повышена, это могло бы компенсировать отмеченные недостатки промежуточных форм, связанные с формированием новой функции. В результате они выстояли бы в борьбе за существование. Приняв, что способность к переключению r- и К-стратегий есть один из механизмов биологической эволюции , мы приходим к вопросу: а как именно оно устроено? Чтобы остаться в пределах представлений об эволюции как закреплении случайно возникших новых признаков путем естественного отбора, нужно принять также, что и переключение стратегий происходит без всякой закономерности, а выживают те, кто выбрал стратегию, более соответствующую данным условиям среды. В простейшем случае должен существовать какой-то один ген или координированная группа генов, режим работы которых и определяет выбор стратегии.
Размножение - производство потомков любым доступным для организма способом.
Биологический смысл
Биологический смысл размножения и связанных с ним процессов довольно разнообразен. Это: по-первых - воспроизводство численности видов и увеличение ее, в противовес естественной смертности, выеданию хищниками и прочим неурядицам. Во-вторых - это обеспечение новых генетических комбинаций и возможность появления новых признаков у потомства, что делает возможным эволюционное развитие группы. Кроме того, в ходе размножения часто решается проблема расселения в пространстве (особенно для малоподвижных видов), переживания периода неблагоприятных условий (чаще всего на стадии покоящихся яиц) и может осуществляться выход на новые пищевые ресурсы (доступные только молоди или личинкам, но не взрослым организмам).
Проблемы и приспособления.
Лишь немногие организмы, главным образом примитивные, способны размножаться пассивно (например, когда их порвут на части). Кроме того, подобный способ (бесполое, оно же вегетативное размножение) не обеспечивает одну из главных функций - появление новых признаков у потомства, которые могли бы дать материал для дальнейшей эволюции. Поэтому, как правило, основным для животных является половое размножение. Для него требуется: развитие специальных систем органов для созревания половых клеток, обеспечение спаривания этих клеток (мужских и женских) от разных особей, обеспечение этих клеток питанием для развития и роста зародыша, а нередко также и дальнейшая забота о молоди до обретения ей самостоятельности.
Возникает как минимум несколько разных сложных моментов. Во-первых, малоподвижные (тем более прикрепленные) организмы должны как-то решить проблему поиска партнера и спаривания (на первый взгляд, практически неразрешимую, особенно при небольшой плотности популяции). Во-вторых, появляющаяся при размножении молодь в любом случае сильно отличается от взрослых особей - она во много раз меньше, что требует выработки новых жизненных стратегий (других механизмов питания , защиты от хищников, осморегуляции и т.п.). Наконец, приходится решать вопросы, связанные с ростом молоди - то есть специально конструировать все структуры, в том числе скелетные, таким образом, чтобы они могли более или менее непрерывно расти, в конечном счете увеличиваясь во много раз. Впрочем, понятно, что все эти проблемы животным удалось успешно решить, и различаются скорее механизмы их решения.
К- и R - стратегии размножения
Стратегии размножения и заботы о потомстве стали предметом одной из общеэкологических теорий - теории R- и K-стратегий. Считается, что все организмы тяготеют к одной из этих двух стратегий размножения. К-стратеги (обычно крупные животные, доминирующие в стабильных местообитаниях и сложившихся сообществах, например - слоны) размножаются медленно и производят немного, но крупных потомков, которых окружают вниманием и заботой. Напротив, R-стратеги (в целом мелкие животные нарушенных местообитаний, например, крысы) плодятся быстро и помногу, но слабо заботятся о потомстве, что сопровождается высокой детской смертностью (взрослая смертность у них также высока). K-стратегия более выгодна в условиях, когда благополучие популяции определяется в основном конкуренцией, а R-стратегия - при сильном влиянии жестких . У человека различные стратегии проявляются даже в пределах вида: в городских популяциях (особенно в экономически развитых странах) люди размножаются медленно (едва обеспечивая воспроизводство), но вкладывают массу средств в содержание, воспитание и образование детей. Напротив, в бедных аграрных странах тропиков люди размножаются быстро и активно, не имея средств прилично одеть, обуть, обучить и иногда даже накормить детей, что нередко приводит к высокой детской смертности, но может и сопровождаться резкими вспышками численности (которые, кстати, отчасти и удерживают низкий уровень жизни в этих странах).
Вся эта теория, впрочем, разработана преимущественно для наземных позвоночных животных (и отчасти для наземных высших растений). В среде водных беспозвоночных действуют несколько иные закономерности. Чаще всего (особенно в море) происходит наоборот - крупные и массовые организмы выбрасывают миллионы микроскопических расселительных яиц или личинок; мелкие же гидробионты расселяются сами, а потомков производят гораздо меньше. Поясним это на примерах.
Сравнительный обзор размножения разных таксонов
Одноклеточные водоросли. В каждой группе одноклеточных водорослей существует два типа размножение - вегетативное и половое. Вегетативное - деление клеток в результате митоза. При обеспеченности ресурсами клетки одноклеточных водорослей размножаются преимущественно вегетативно, и численность популяции возрастает по экспоненте. При неблагоприятных для вегетативного деления условиях или в результате других причин у водорослей происходит половое размножение (мейоз), при котором образуются мужские и женские гаметы, после слияния которых формируется клетка с «новым» генотипом. Жизненные циклы одноклеточных водорослей, принадлежащих к разным филогенетическим группам, различаются. Циклы многих водорослей включают покоящиеся стадии - (покоящиеся клетки, споры, цисты и др.) для переживания неблагоприятных условий.
Беспозвоночные. Исходный (для водных, в первую очередь морских беспозвоночных) тип размножения, как считается, выглядит следующим образом. Примерно в одно время все взрослые мужские и женские особи в большом количестве выметывают прямо в воду свои половые продукты (яйцеклетки и сперматозоиды), которые сами (если повезет) находят друг друга в толще воды и спариваются. Это называется наружним оплодотворением. Сам организм при этом может быть малоподвижным или сидячим. Из оплодотворенной зиготы вырастает микроскопическая планктонная личинка, которая довольно длительное время плавает в толще воды, расселяясь с течениями, претерпевая разнообразные превращения и со временем переходя на внешнее питание (чаще всего фитопланктоном - так называемая планктотрофная личинка). Подрастая и готовясь переходить ко взрослому образу жизни, личинка оседает на подходящий донный субстрат и приобретает признаки взрослой особи, достигает макроскопических размеров и в дальнейшем еще долго растет. Этот тип размножения и развития позволяет решать все проблемы расселения и внутривидовой конкуренции именно за счет личинок (а взрослые особи могут быть даже сидячими - им не нужно непосредственно встречаться друг с другом). С другой стороны, такой подход сопровождается огромной смертностью как среди гамет, так и среди личинок, что требует массового их накопления и выброса, причем крайне важна синхронизация созревания и выброса половых клеток у разных особей популяции. Это достигается выбросом в воду сигнальных веществ, стимулирующим у особей выброс в воду всех накопленных заранее гамет. Обычно массовый нерест происходит один раз в год, причем у многих организмов - один раз в жизни. Как легко понять, такая стратегия удобна для относительно крупных, массовых, массивных и малоподвижных организмов: полипов, губок, моллюсков, крупных полихет, иглокожих и ракообразных. В общем, в море этот вариант считается самым типичным.
А также мелкие и подвижные беспозвоночные (ветвистоусые и веслоногие ракообразные, некоторые мелкие полихеты, олигохеты, улитки) не могут позволить себе массовый выброс гамет в воду (просто не имея достаточно массы), и применяют внутреннее оплодотворение: находят друг друга и спариваются сами, после чего самка, как правило, еще некоторое время вынашивает развивающиеся яйца внутри себя (уменьшая их смертность). Рождаются либо защищенные специальной оболочкой пассивные яйца, либо уже активные личинки. Личинки чаще всего ведут образ жизни, сходный со взрослыми; но нередко более подвижны, что обеспечивает популяции лучшее расселение в пространстве. Иногда и в этом случае личинки бентосных организмов на некоторое время становятся планктонными. Часто (например, у олигохет), личинок вообще нет, а молодь по строению и образу жизни сходна со взрослыми (прямое развитие). Все это позволяет генерировать на несколько порядков меньше половых продуктов, снижая репродуктивные затраты, и при этом размножаться круглый год, не заботясь о синхронизации нереста. Часто личинка при рождении снабжается запасом питательных веществ, достаточным для прохождения всей ее личиночной расселительной жизни, и вообще не питается (лецитотрофная личинка).
В пресных водах размножение по первому типу (с наружним оплодотворением и длительной стадией планктонной личинки) затруднено осмотическими проблемами: осморегулировать плавающие гаметы и планктонных личинок оказалось крайне неудобным, и большинство даже низших беспозвоночных применяют внутреннее оплодотворение - и никаких дополнительных планктонных личинок. Как правило, откладываются достаточно крупные яйца - в небольшом количестве, но с приличным запасом питательных веществ, что позволяет организму быть в большой степени лецитотрофным и вылупляться, будучи уже вполне макроскопическим и с развитой системой осморегуляции. Это путь пресноводных червей, улиток и большинства ракообразных. Веслоногие рачки (вроде циклопов) все же имеют планктонную личинку (науплиуса), но относительно короткоживущую, в серии последовательных линек быстро достигающую дефинитивного (взрослого) облика.
Насекомые, как группа в целом наземная, причем высокоподвижная именно на стадии взрослой особи (имаго), при освоении водной среды выработали свою стратегию размножения и жизненного цикла. Они оставили на долю взрослых особей именно расселительную функцию (а также спаривание и откладку яиц), а личинки, обитающие в воде (причем обычно гораздо дольше, чем имаго) отвечают за питание, рост (и накопление питательных веществ в теле), а также переживание в воде сезонов, неблагоприятных для жизни на суше (главным образом зимы). Личинки насекомых уже при вылуплении из яиц макроскопичны, способны к самостоятельному питанию и имеют вполне совершенную систему пресноводной (а иногда и солоноватоводной) осморегуляции. Интересно, что имаго у некоторых групп (поденки, ручейники, хирономиды, часть веснянок) вообще не питаются и живут очень недолго, и для успешного размножения применяется их синхронизированный вылет из водоемов. Таким образом, взрослые насекомые у несекомых приравниваются, функционально, к половым продуктам (гаметам) у многих морских беспозвоночных.
В некоторых группах беспозвоночных (чаще в пресных водах, чем в морских) распространен гермафродитизм - когда в каждой особи формируются и мужские, и женские половые органы и гаметы. Например, гермафродиты все легочные улитки (Pulmonata), олигохеты, усоногие ракообразные. При спаривании организм может выступать и самцом, и самкой, а часто - обоими сразу (тогда наблюдается взаимное оплодотворение). Биологический смысл гермафродитизма (то есть выращивания в каждом теле двойного набора органов) не вполне ясен. Иногда (но, видимо, редко) происходит самооплодотворение - это отчасти нарушает саму идею полового размножения (поскольку организм скрещивается сам с собой), зато позволяет единичной особи породить на новом месте новую популяцию.
Еще реже, чем гермафродитизм, у животных наблюдается бесполое размножение, при котором материнские особи фактически клонируют себя, порождая генетически точно такие же женские особи. Такая ситуация особенно характерна для периодов вспышек численности мелких пресноводных беспозвоночных - в частности, дафний и коловраток в летний период. В любом случае - это временная мера, рано или поздно (обычно осенью) сменяющаяся нормальным половым размножением. Впрочем, у одноклеточных простейших (как и у растений) бесполое размножение - самая обычная вещь, именно за счет его происходит основное воспроизведение видов.
Рыбы. Как правило, рыбы имеют наружнее оплодотворение, однако осуществляемое при личной встрече родительских особей (самка откладывает свои яйцеклетки, а самец тут же поливает ее молоками). Соответственно, рыбы мечут икру, обычно довольно мелкую и в больших количествах. Число икринок в среднем составляет несколько тысяч, но сильно варьирует у разных видов: от 10-30 штук (у колюшек) до 10-100 миллионов (у тунцов, трески и многих других крупных морских рыб). При этом икринки несут некоторый запас питательных веществ, что позволяет вылупляться из икры уже вполне сформировавшимся малькам, способным к плаванию и питанию. Никаких новых сред мальки рыб не осваивают, зато перехватывают спектры питания, обычно недоступные взрослым рыбам: они могут питаться зоопланктоном и мейобентосом. Правда, не ясно, сильно ли выигрывают от этого обстоятельства рыбы в целом или это вынужденная мера (поскольку ничем другим мальки рыб питаться не в состоянии за малостью размеров).
Отдельные виды рыб, тем не менее, имеют довольно причудливые формы размножения и защиты потомства. Наиболее известны проходные рыбы, меняющие ради размножения среду обитания. Лососевые и осетровые во взрослом состоянии живут в морях, а нерестятся в реках (где и произошли), и молодь их, приспособленная к пресноводной осморегуляции, некоторое время держится в реках и лишь потом спускается в море. При этом лососи показывают чудеса героизма, преодолевая пороги горно-таежных рек; а вскоре после нереста отмирают - прямо в реках, резко увеличивая их сапробность . Получается, что это такой своеобразный способ насытить органикой местообитание молоди. Другой вопрос, насколько он эффективен.
Угорь, напротив, плывет на размножение из рек в Саргассово море, причем его европейская популяция ради этого преодолевает (по течению) почти весь Атлантический океан. Молодь (опять же по течению, но уже по другому) возвращается к европейским рекам. Большого смысла в этом, кажется, нет. Считается, что сверхдлинные миграции угря отражают дрейф континентов, в ходе которого Атлантика постепенно расширяется, и угрям с каждым миллионом лет приходится плавать в родное море все дальше и дальше.
Некоторые группы рыб перешли к явной K-стратегии, главным образом путем живорождения. При этом оплодотворение внутреннее, число потомков гораздо меньше, но сами они в момент выхода в воду крупнее и жизнеспособнее. Наиболее известный пример - живородящие аквариумные рыбки Peciliidae (гуппи и прочие пецилии). Все аквариумисты знают, что развести их гораздо проще, чем любых других рыб. Сходным образом поступают, к примеру, акулы - они откладывают очень мало икринок (обычно 5-30), но очень крупных - у китовой акулы до 60 см (!) в диаметре, что позволяет вылупляться из них уже весьма крупным рыбам.
Земноводные . Амфибии имеют внутреннее оплодотворение и откладывают довольно крупную икру - причем обязательно в воду. Подобно насекомым, большинство земноводных амфибиотичны - то есть размножаются именно в воде и имеют водных рыбообразных личинок (головастиков), хотя взрослые животные большую часть жизни обитают на суше. В общем, здесь также можно говорить о перехвате головастиками новой среды обитания и пищевых ресурсов - это в общем верно, но на самом деле отражает глобальную неполноценность всего класса - земноводные просто не умеют иначе.
Крабы и забота о женщине . У многих ракообразных, особенно высших, взрослые особи столь надежно защищены хитиновым панцирем, что не могут спариваться, кроме как сразу после линьки у самки. Поэтому готовый к спариванию самец должен не просто найти самку своего вида, но и дождаться ее линьки, что может произойти, например, через несколько недель. Причем нужно именно ждать рядом, а не искать линяющую самку - поскольку при линьке и после нее животные становятся крайне уязвимы и стараются линять в надежных убежищах (где их трудно найти). Поэтому, например, у камчатских крабов взрослые самцы собирают вокруг себя несколько самок (гарем) и «пасут» их, спариваясь с теми, кто полинял, и их же обороняя от поедания (в первую очередь другими самками своего же гарема). Такая жизненно необходимая забота о самке имеет мало общего с легкомысленным «ухаживанием» позвоночных. Ситуация осложняется еще и тем, что в случае линьки самого самца он также может быть немедленно съеден своими самками, поэтому для линьки он вынужден покидать свой гарем и тщательно прятаться.
Гарпактициды и педофилия . Эти мелкие веслоногие рачки обладают слабым половым диморфизмом, да и межвидовые различия у них невелики; и возрастные изменения (от копеподитных стадий молоди к половозрелым) слабо заметны. Зато инстинкт спаривания у самцов весьма силен. Поэтому готовый к спариванию самец, роясь в донном наилке в поисках полового партнера, не проявляет разборчивости и спаривается практически с кем угодно - с самкой своего вида (если повезет!), или с самцом, или с копеподитом (то есть молодой особью), или с рачком совсем другого вида. Иногда их подобную активность принимают за попытку съесть партнера, но это попытка копуляции. В состоянии спаривания рачки плавают довольно долго, причем если в позиции самки также оказывается самец, он может тем временем также ловить себе партнера для спаривания; иногда таким образом получаются довольно длинные цепочки особей, лишь немногие из которых действительно спариваются.
Улитки и групповое спаривание . Легочные пресноводные улитки - гермафродиты, причем у некоторых из них определение пола при спаривании прямо определяется позицией самого животного - приблизительно по принципу «кто сверху, тот и самец». Например, речные чашечки (Ancylus fluviatilis ) для спаривания просто наползают одна на другую, а затем свешивают вниз совокупительные органы. Эта ситуация не мешает еще одной чашечке заползти сверху и копулировать с нижесидящей, и так далее. В итоге может образоваться стопка копулирующих особей, из которых самая нижняя выступает лишь как самка, а самая верхняя - как самец, а все остальные работают обоими системами органов (в отличие от глупых гарпактицид, которые могут лишь имитировать подобную ситуацию). Потом все расползаются и дружно откладывают яйца.
Бонеллия и определение пола судьбой . У сидячей морской эхиуриды бонеллии планктонная личинка, выходя в открытое плавание, еще не имеет определенного пола, но уже имеет не только расселительную, но и половую задачу - поиск самки. Если личинке удается найти взрослую самку бонеллии, она проникает в нее и развивается в самца (который всю жизнь потом живет внутри самки, оплодотворяя ее). Если же найти самку так и не удается, личинка в конце концов оседает на дно и сама становится самкой.
История развития понятия «экологическая стратегия » у растений .
В - первые, термин «стратегия» означал совокупность свойств, помогающих организмам выживать в данных условиях, и применялся только по отношению к животным организмам.
Выделялись R- и К-стратегии по соотношению затрат на размножение и поддержание потомства.
К-стратеги отличаются заботой о немногочисленном потомстве, это наблюдается, например, у слонов. R-стратеги характеризуются максимальной плодовитостью и отсутствием заботы о потомстве, например, аскариды.
Свойства К- и R -стратегий у животных.
| R -стратегия | К-стратегия |
| Характерно быстрое развитие особей | Характерно медленное развитие особей |
| Высокая плодовитость | Низкая плодовитость |
| Мелкие размеры особей | Крупные размеры особей |
| Короткая продолжительность жизни | Значительная продолжительность жизни |
| Более ранние акты размножения | Позднее размножение |
| Все признаки направлены на более высокую продуктивность | Все признаки направлены на болнн эффективное использование ресурсов |
| Характерна для катастрофических изменений среды, при заселении незаполненных биотопов. | Наиболее эффективна в конкурентной среде. |
Позже термин «экологическая стратегия» стал использоваться и применительно к растительным организмам. (20).
Для отечественной литературы термин « стратегия» применительно к растениям достаточно нов и первым его использовал Т.А. Работнов (1975 г.), назвавший так выделенные Л.Г. Раменским (1936 г.) «ценобиотические типы».
Под стратегией вида Работнов предложил понимать «совокупность приспособлений, обеспечивающих ему возможность обитать совместно с другими организмами и занимать определенное место в соответствующем биогеоценозе». (10)
Первым на наличие предпосылок у растений, определяющих их статус в сообществе, еще в 1894 г. обратил внимание Мак Леод, который разделил все виды на « капиталистов» и «пролетариев».
Однако малоудачной была как сама аналогия с обществом, так и основной критерий различения типов_ перекрестное опыление и самоопыление, хотя ученый пытался сделать оценки комплексным и писал о том, что «капиталистам» свойственны наличие запаса питательных веществ, поликарпичность, нетерпимость к затенению и т.д.
Блестящую разработку этот вопрос получил в работах Раменского, опубликованных в 30-ых годах, где он писал о 3-х типах растений, которые назвал виолентами, патиентами и эксплерентами и уподобил их львам, верблюдам и шакалам.
Спустя 40 лет в Англии вышла монография Дж. Грайма «Стратегии растений и процессы в растительности.» , в которой автор, не зная работ Раменского, заново описывал те же три типа стратегий под названием конкурентов, стресс-толерантов и рудералов.
Для понимания типа стратегий многое было сделано также Э. Пианкой, Р. Уиттекером и Т.А. Работновым. (11)
Основные системы эколого-ценотических стратегий .
Система Э. Пианки .
Система Пианки, получившая самое широкое распространение в экологии, включает два типа стратегий, связанных с К-отборами и r-отборами (по соотношению долей энергетических затрат на поддержание взрослых особей и на процессы размножения).
К- отбор – это отбор в постоянной (предсказуемой) среде, где основная часть энергии популяции затрачивается на конкуренцию, а при r –отборе- основной статьей расхода энергии является размножение.
Система явилась результатом развития представлений, которые были сформулированы ранее Р.Х. Макартуром и Е.О. Уилсоном, однако именно Э. Пианка всесторонне проанализировал те следствия, которые возникают в результате реализации двух типов отбора.
Два типа стратегии Пианки имеют в мире растений самое широкое распространение. И даже возникновение разноспоровости у плаунов или папоротников можно рассматривать в конечном итоге как замену r-стратегии изоспор на К-стратегию женского гаметофита, которая гарантирует лучшее выживание потомства и заменяет огромное количество мелких изоспор ограниченным числом мегаспор, обеспечивающих необходимые условия развития женского заростка.
К-стратеги приурочены к более или менее стабильным условиям среды, обладают равновесными популяциями, где смертность регулируется плотностью, приспособлены к условиям острой конкуренции. Они, как правило, поликарпики с медленным развитием и жизненной формой от трав до деревьев. В сукцессионных сериях эти виды увеличивают свое участие по мере приближения сукцессионной стадии к климаксу.
r-стратеги, напротив, предпочитают нестабильные местообитания, характеризующиеся неравновесными популяциями, смертность которых не зависит или зависит в слабой степени от плотности. Конкуренция между такими растениями слабая, это малолетники-монокарпики, как правило, травы, реже - кустарники. В сукцессионном ряду они связаны с пионерными стадиями и не играют существенной роли в зрелых сообществах, предваряющих климакс.
Таким образом, система типов Э. Пианки проста – одномерна, однако она полностью соответствует континуальному восприятию типов.
Он отмечает относительность разделения всех видов на 2 типа стратегий, подчеркивая, что мир не окрашен только в черное и белое, и крайние варианты, как правило, связаны целой гаммой переходов (Э. Пианка, 1981 г., стр. 138). (13)
Система Р. Уиттекера .
Р. Уиттекер (1980) различал не 2, а три типа стратегий, обозначаемые буквами К,r и L. В основу его системы положены закономерности колебания численности популяций между двумя пределами: К-верхний предел, соответствующий максимальной плотности насыщения и L- нижний предел, означающий некий « популяционный ноль», соответствующий численности, которая не способна обеспечить выживание популяции.
К-стратеги стремятся к достижению уровня К, добиваясь этого, во-первых, за счет предельной дифференциации ниш. К-отбор воздействует на механизмы, с помощью которых сохраняют свою популяцию в процессе конкуренции и других взаимодействий в границах занятой ими среды. Численность популяций существенно снижается, однако общей тенденцией таких популяций является колебания вокруг уровня К.
Вторая группа популяций_r-стратеги. Они характеризуются резкими флюктуациями между уровнями К и L. Такие популяции нестабильны и выживают лишь благодаря высокой скорости продуцирования диаспор, они слабо адоптированы как к условиям обостренной конкуренции, так и к неблагоприятным условиям, вызывающим стресс.
Третья группа популяций-L-стратеги, которые флюктуцируют около нижнего предела численности L, хотя могут временами взрывообразно увеличивать свою численность. У таких популяций отбор имеет тенденцию к совершенствованию механизма для переживания неблагоприятных периодов, а скорость размножения может быть или не быть высокой.
Различая три типа отбора с их результатом – тремя первичными типами в то же время Уиттекер, как и Пианка, не абсолютировал своей системы.
Если сравнивать системы Уиттекера и Пианки, очевидно, что его типы К и r соответствуют К и r Пианки и действительно дифференциация ниш идет под действием К-отбора. Это в основном многолетние виды, часто размножающиеся вегетативно, и расходующие в генеративной сфере сравнительно мало энергии.
Рудеральные растения, напротив, отличаются укороченностью жизненного цикла и высокой семенной продуктивностью, и поэтому затраты на размножение здесь больше. Это является следствием r-отбора.
Группа L занимает переходное положение, так как пустынные однолетники относятся к числу эфемеров с очень быстрым циклом развития и высокой семенной продуктивностью (результат r-отбора), но кустарнички, а также некоторые травянистые дерновинные растения переживают стресс в вегетативном состоянии и потому представляют результат действия К-отбора. (10)
Система Раменского-Грайма.
Раменский предложил систему из трех типов. Он различал три «ценобиотических типа».
Первый тип, который он называл виолентами или «львами», характеризуется способностью к энергичному захвату территории, полнотой используемых ресурсов, мощным конкурентным подавлением соперников.
Второй тип - патиенты или «верблюды» отличаются способностью к перенесению экстремальных условий среды, то есть выносливостью.
Третий тип – эксплеренты или шакалы не отличаются ни устойчивостью к стрессовым ситуациям, ни высокой конкурентной мощностью, но способны к быстрому захвату промежутков между более сильными растениями, и при их смыкании также легко вытесняются. (13)
В дальнейшем представления и классификация Л.Г. Раменского (1935-38 гг.) развивались Работновым Т.А. (1966, 1975, 1978, 1980 гг.). Им показан сложный характер патиентности (стресс-толерантности) у растений и выделены экологические и фитоценотические патиенты.
Первые способны существовать в неблагоприятных условиях за счет экологической специализации (на засоленных, кислых, сухих или каменистых субстратах и т.д.) и в наибольшей степени соответствуют патиентам Л.Г. Раменского. У них совпадают и аутэкологические и синэкологические оптимумы.
Вторые способны длительно выживать под прессом виолентов в экологически оптимальных условиях при помощи максимального снижения процессов жизнедеятельности. Синэкологические и аутэкологические оптимумы у них обычно не совпадают. (6 )
Дальнейшее развитие представлений о типах стратегий мы находим в многочисленных работах Дж. Грайма (J.Grime, 1974, 1978, 1979).
Он предлагает, по существу, 3, таких же как и у Л.Г. Раменского, типа эколого-ценотических стратегий, назвав эти типы: конкурентами, стресс толерантами и рудералами (соответственно К, S и R).
Экологические стратегии популяций
Каковы бы не были приспособления особей к совместному проживанию в популяции, каковы бы не были приспособления популяции к тем или иным факторам, все они в конечном итоге направлены на долгое выживание и продолжение себя в любых условиях существования. Среди всех приспособлений и особенностей можно выделить комплекс базовых признаков, которые называются экологической стратегией. Это общая характеристика роста и размножения данного вида, включающая темп роста особей, период достижения ими половой зрелости, периодичность размножения, предельный возраст и пр.
Экологические стратегии очень разнообразны и хотя между ними существует множество переходов, из них можно выделить два крайних типа: r-стратегию и K-стратегию.
r-стратегия – ею обладают быстро размножающиеся виды (r-виды); для нее характерен отбор на повышение скорости роста популяции в периоды низкой плотности. Она характерна для популяций в среде с резкими и непредсказуемыми изменениями условий или в эфемерных, ᴛ.ᴇ. существующих короткое время (пересыхающие лужи, заливные луга, временные водотоки)
Основные признаки r-видов: высокая плодовитость, короткое время регенерации, высокая численность, обычно малые размеры особей (у растений мелкие семена), малая продолжительность жизни, большие траты энергии на размножение, кратковременность местообитаний, низкая конкурентоспособность. R-виды быстро и в больших количествах заселяют не занятые территории, но, как правило, скоро – в течение жизни одного-двух поколений сменяются К-видами.
К r-видам относятся бактерии, все однолетние растения (сорняки) и насекомые-вредители (тли, листоеды, стволовые вредители, стадная фаза саранчи). Из многолетников – пионерные виды: Иван-чай, многие злаки, полыни, эфемерные растения, из древесных видов – ивы, березы белая и каменная, осина, чозения, из хвойных – лиственница; они появляются первыми на нарушенных землях: гарях, горных полигонах, строительных карьерах, по обочинам дороᴦ.
K-стратегия – этой стратегией обладают виды с низкой скоростью размножения и высокой выживаемостью (К-виды); она определяет отбор на повышение выживаемости при высокой плотности популяции, приближающейся к предельной.
Основные признаки К-видов: низкая плодовитость, значительная продолжительность жизни, крупные размеры особей и семян, мощные корневые системы, высокая конкурентоспособность, устойчивость на занимаемой территории, высокая специализация образа жизни. Скорость размножения К-видов с приближением к предельной плотности популяции падает и быстро увеличивается при низкой плотности; родители заботятся о потомках. К-виды часто становятся доминантами биогеоценозов.
К К-видам относятся все хищники, человек, реликтовые насекомые (крупные тропические бабочки, в т.ч. дальневосточные, реликтовый усач, жук-олень, жужелицы и др.), одиночная фаза саранчи, почти все деревья и кустарники. Наиболее яркие представители растений – все хвойные, дуб монгольский, орех маньчжурский, лещины, клены, разнотравье, осоки.
Разные популяции по-разному используют одну и ту же среду обитания, в связи с этим в ней одновременно могут существовать виды обоих типов стратегией.
ПРИМЕРЫ. В лесах на экологическом профиле "Горнотаежный" весной до распускания листьев на деревьях, спешат зацвести, отплодоносить и закончить вегетацию эфемероиды: хохлатки, адонис амурский, ветреницы, фиалка восточная (желтая). Под пологом леса начинается цветение пионов, лилий, воронца остроконечного. На открытых участках в сухих дубняках южного склона разрастаются овсяница овечья и марьянник розовый. Дуб, овсяница и другие виды – K-стратеги, марьянник – r-стратеᴦ. 40 лет назад после пожара в пихтово-широколиственном типе леса образовались парцеллы из осины (r-вид). Сегодня осина уходит из состава древостоя, сменяясь К-видами: липой, дубом, грабом, орехом и др.
Любая популяция растений, животных и микроорганизмов - ϶ᴛᴏ совершенная живая система, способная к саморегуляции, восстановлению своего динамического равновесия. Но она существует не изолированно, а совместно с популяциями других видов, образуя биоценозы. По этой причине в природе широко распространены и межпопуляционные механизмы, регулирующие взаимоотношения между популяциями разных видов. В качестве регулятора данных взаимоотношений выступает биогеоценоз, состоящий из множества популяций разных видов. В каждой из этих популяций происходят взаимодействия между особями, и каждая популяция оказывает воздействие на другие популяции и на биогеоценоз в целом, как и биогеоценоз с входящими в него популяциями оказывает непосредственное влияние на каждую конкретную популяцию.
Как пишет И.И. Шмальгаузен: "…Во всех биологических системах имеется всегда взаимодействие разных циклов регуляции, ведущее к саморазвитию системы соответственно данным условиям существования…"
При достижении оптимальных соотношений наступает более или менее долгое стационарное состояние (динамическое равновесие) данной системы в данных условиях существования. "…Для популяции это означает установление определенной генетической структуры, в том числе, разных форм сбалансированного полиморфизма. Для вида это означает установление и поддержание его более или менее сложного строения. … Для биогеоценоза это означает установление и поддержание его гетерогенного состава и сложившихся соотношений между компонентами. При изменении условий существования стационарное состояние, конечно, нарушается. Происходит переоценка нормы и вариантов, а, следовательно, и новое преобразование, ᴛ.ᴇ. дальнейшее саморазвитие данных систем…". При этом в биогеоценозе изменяются соотношения между звеньями, а в популяциях идет перестройка генетической структуры.
Экологические стратегии популяций - понятие и виды. Классификация и особенности категории "Экологические стратегии популяций" 2017, 2018.